RéférencesBerardi, N., Pizzorusso, T., Ratto, G. M., & Maffei, L. (2003). Molecular basis of plasticity in the visual cortex. Trends Neurosci, 26(7), 369-378.
Bittar, R.G., Olivier, A., Sadikot, A.F., Anderman, F., Pike, G.B., Reutens, D.C. (1999) Presurgical motor and somatosensory cortex mapping with functional magnetic resonance imagery and positron emission tomography. J Neursurg, 91(6)915-921
Blakemore, C., Van Sluyters, C. V., & Movshon, J. A. (1976). Synaptic competition in the kitten's visual cortex. Cold Spring Harb Symp Quant Biol, 40, 601-609.
Boire, D., Dufour, J. S., Theoret, H., & Ptito, M. (2001). Quantitative analysis of the retinal ganglion cell layer in the ostrich, Struthio camelus. Brain Behav Evol, 58(6), 343-355.
Boire, D., Theoret, H., Herbin, M., Casanova, C., & Ptito, M. (2000) Retinogeniculate projections following early cerebral hemispherectomy in the vervet monkey. Exp Brain Res, 135(3): 373-381.
Bronchti, G., Heil, P., Sadka, R., Hess, A., Scheich, H., & Wollberg, Z. (2002). Auditory activation of "visual" cortical areas in the blind mole rat (Spalax ehrenbergi). Eur J Neurosci, 16(2), 311-329.
Chebat, D. R., Boire, D., & Ptito, M. (2006)a. Development of the commissure of the superior colliculus in the hamster. J Comp Neurol, 494(6), 887-902.t
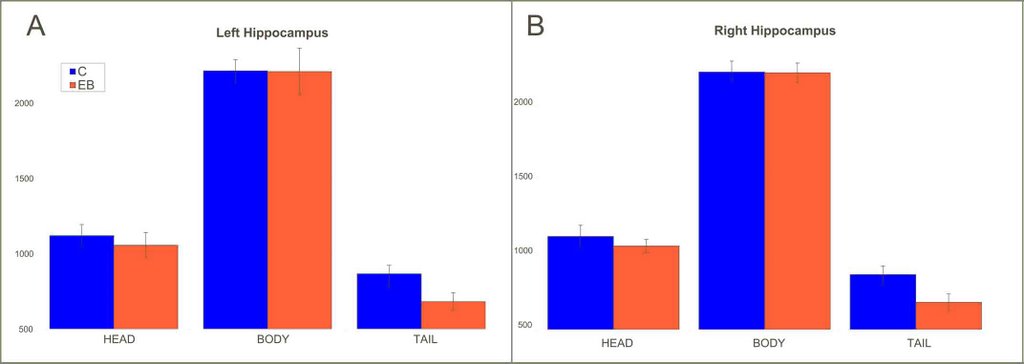
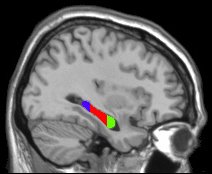
Chebat, D.R., Chen, J.K., Ptito., A., Schneider, F., Kupers, R., Ptito, M. (2006) Volumetric anlysis of the hippocmapus in early nlind subjects. OHBM , Florence, Italy.
Constantine-Paton, M., & Law, M. I. (1978). Eye-specific termination bands in tecta of three-eyed frogs. Science, 202(4368), 639-641.
Crane, J., & Milner, B. (2005). What went where? Impaired object-location learning in patients with right hippocampal lesions. Hippocampus, 15(2), 216-231.
Crowley, J. C., & Katz, L. C. (1999). Development of ocular dominance columns in the absence of retinal input. Nat Neurosci, 2(12), 1125-1130.
Crowley, J. C., & Katz, L. C. (2000). Early development of ocular dominance columns. Science, 290(5495), 1321-1324.
De Volder, A. G., Bol, A., Blin, J., Robert, A., Arno, P., Grandin, C., Michel, C., & Veraart, C. (1997). Brain energy metabolism in early blind subjects: neural activity in the visual cortex. Brain Res, 750(1-2), 235-244.
Dehay, C., Giroud, P., Berland, M., Killackey, H., & Kennedy, H. (1996a). Contribution of thalamic input to the specification of cytoarchitectonic cortical fields in the primate: effects of bilateral enucleation in the fetal monkey on the boundaries, dimensions, and gyrification of striate and extrastriate cortex. J Comp Neurol, 367(1), 70-89.
Dehay, C., Giroud, P., Berland, M., Killackey, H. P., & Kennedy, H. (1996b). Phenotypic characterisation of respecified visual cortex subsequent to prenatal enucleation in the monkey: development of acetylcholinesterase and cytochrome oxidase patterns. J Comp Neurol, 376(3), 386-402.
Feldman, D. E., & Brecht, M. (2005). Map plasticity in somatosensory cortex. Science, 310(5749), 810-815.
Fendrich, R., Wessinger, C.M., & Gazzaniga, M.S. (2001) Speculations on the neural basis of islands of blindsight. Prog Brain Res, 134: 353-366
Frost, D. O. (1982). Anomalous visual connections to somatosensory and auditory systems following brain lesions in early life. Brain Res, 255(4), 627-635.
Frost, D. O., Boire, D., Gingras, G., & Ptito, M. (2000). Surgically created neural pathways mediate visual pattern discrimination. Proc Natl Acad Sci U S A, 97(20), 11068-11073.
Frost, D. O., & Metin, C. (1985). Induction of functional retinal projections to the somatosensory system. Nature, 317(6033), 162-164.
Guillery, R. W., & Stelzner, D. J. (1970). The differential effects of unilateral lid closure upon the monocular and binocular segments of the dorsal lateral geniculate nucleus in the cat. J Comp Neurol, 139(4), 413-421.
HEBB, C. O., & KONZETT, H. (1949). The effect of certain analgesic drugs on synaptic transmission as observed in the perfused superior cervical ganglion of the cat. Q J Exp Physiol Cogn Med Sci, 35(3), 213-217.
Holdstock, J. S., Mayes, A. R., Cezayirli, E., Isaac, C. L., Aggleton, J. P., & Roberts, N. (2000). A comparison of egocentric and allocentric spatial memory in a patient with selective hippocampal damage. Neuropsychologia, 38(4), 410-425.
Innocenti, G. M. (1978). Postnatal development of interhemispheric connections of the cat visual cortex. Arch Ital Biol, 116(3-4), 463-470.
Innocenti, G. M., & Frost, D. O. (1980). The postnatal development of visual callosal connections in the absence of visual experience or of the eyes. Exp Brain Res, 39(4), 365-375.
Kandel, E. R. (2001). The molecular biology of memory storage: a dialogue between genes and synapses. Science, 294(5544), 1030-1038.
Korte, M., & Rauschecker, J. P. (1993). Auditory spatial tuning of cortical neurons is sharpened in cats with early blindness. J Neurophysiol, 70(4), 1717-1721.
Kupers, R., Fumal, A., Schonen, F., & Ptito, M., (2005) Tactile phosphene elicited by TMS in the blind: new evidence for the development of cross modal plasticity. OHBM 2005, Toronto, Canada.
Law, M. I., & Constantine-Paton, M. (1980). Right and left eye bands in frogs with unilateral tectal ablations. Proc Natl Acad Sci U S A, 77(4), 2314-2318.
Law, M. I., & Constantine-Paton, M. (1981). Anatomy and physiology of experimentally produced striped tecta. J Neurosci, 1(7), 741-759.
Leh, S. E., Johansen-Berg, H., & Ptito, A. (2006). Unconscious vision: new insights into the neuronal correlate of blindsight using diffusion tractography. Brain, 129(Pt 7), 1822-1832.
Maguire, E. A. (1997). Hippocampal involvement in human topographical memory: evidence from functional imaging. Philos Trans R Soc Lond B Biol Sci, 352(1360), 1475-80.
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak, R. S., & Frith, C. D. (2000). Navigation-related structural change in the hippocampi of taxi drivers. Proc Natl Acad Sci U S A, 97(8), 4398-403.
Maguire, E. A., Mummery, C. J., & Buchel, C. (2000). Patterns of hippocampal-cortical interaction dissociate temporal lobe memory subsystems. Hippocampus, 10(4), 475-82.
Mellet, E., Briscogne, S., Tzourio-Mazoyer, N., Ghaem, O., Petit, L., Zago, L., Etard, O., Berthoz, A., Mazoyer, B., & Denis, M. (2000). Neural correlates of topographic mental exploration: the impact of route versus survey perspective learning. Neuroimage, 12(5), 588-600.
Miller, B., Windrem, M. S., & Finlay, B. L. (1991). Thalamic ablations and neocortical development: alterations in thalamic and callosal connectivity. Cereb Cortex, 1(3), 241-61.
O'Keefe, J. (1991). An allocentric spatial model for the hippocampal cognitive map. Hippocampus, 1(3), 230-5.
O'Keefe, J., & Speakman, A. (1987). Single unit activity in the rat hippocampus during a spatial memory task. Exp Brain Res, 68(1), 1-27.
Olausson, H., Marchand, S., Bittar, R.G., Bernier, J., Ptito, A., Bushnell, M.C. (2001) Central Pain in a hemispherctomized patient. Eur J Pain, 5(2) : 209-217
Olavarria, J. F., & Hiroi, R. (2003). Retinal influences specify cortico-cortical maps by postnatal day six in rats and mice. J Comp Neurol, 459(2), 156-72.
Ptito, A., Fortin, A., & Ptito, M. (2001) `Seeing`in the blind hemifield following hemispherectomy. Prog Brain Res, 134: 367-378
Ptito, M., Bouchard, P., Lepore, F., Quessy, S., Di Stefano, M., & Guillemot, J. P. (1995). Binocular interactions and visual acuity loss in esotropic cats. Can J Physiol Pharmacol, 73(9), 1398-405.
Ptito, M., Dalby, M., Gjedde, A., (1999) Visual field recovery in a patient with bilateral occipital lobe damage. Acta Neurol Scand, 99(4): 252-254.
Ptito, M., Desgent, S., Sensory input-based adaptation and brain architecture (2006) Lifespan development and the brain. Baltes, New-York press.
Ptito, M., Giguere, J. F., Boire, D., Frost, D. O., & Casanova, C. (2001). When the auditory cortex turns visual. Prog Brain Res, 134, 447-58.
Ptito, M., & Kupers, R. (2005). Cross-modal plasticity in early blindness. J Integr Neurosci, 4(4), 479-88.
Ptito, M., Moesgaard, S. M., Gjedde, A., & Kupers, R. (2005). Cross-modal plasticity revealed by electrotactile stimulation of the tongue in the congenitally blind. Brain, 128(Pt 3), 606-14.
Rakic, P., Suner, I., & Williams, R. W. (1991). A novel cytoarchitectonic area induced experimentally within the primate visual cortex. Proc Natl Acad Sci U S A, 88(6), 2083-7.
Rampon, C., & Tsien, J. Z. (2000). Genetic analysis of learning behavior-induced structural plasticity. Hippocampus, 10(5), 605-9.
Rauschecker, J. P., & Kniepert, U. (1994). Auditory localization behaviour in visually deprived cats. Eur J Neurosci, 6(1), 149-60.
Rauschecker, J. P., & Korte, M. (1993). Auditory compensation for early blindness in cat cerebral cortex. J Neurosci, 13(10), 4538-48.
Rebillard, G., Carlier, E., & Pujol, R. (1977). [Visual evoked responses on the primary auditory cortex in the cat after an early suppression of cochlear receptors (author's transl)]. Rev Electroencephalogr Neurophysiol Clin, 7(3), 284-9.
Rebillard, G., Carlier, E., Rebillard, M., & Pujol, R. (1977). Enhancement of visual responses on the primary auditory cortex of the cat after an early destruction of cochlear receptors. Brain Res, 129(1), 162-4.
Reh, T. A., & Constantine-Paton, M. (1985). Eye-specific segregation requires neural activity in three-eyed Rana pipiens. J Neurosci, 5(5), 1132-43.
Rhoades, R. W., & Fish, S. E. (1982). Altered organization of intercollicular pathway in bilaterally enucleated hamsters. Brain Res, 256(3), 356-60.
Rosa, M. G., Schmid, L. M., & Calford, M. B. (1995). Responsiveness of cat area 17 after monocular inactivation: limitation of topographic plasticity in adult cortex. J Physiol, 482 ( Pt 3), 589-608.
Rubenstein, J. L., Anderson, S., Shi, L., Miyashita-Lin, E., Bulfone, A., & Hevner, R. (1999). Genetic control of cortical regionalization and connectivity. Cereb Cortex, 9(6), 524-32.
Sadato, N., Okada, T., Honda, M., & Yonekura, Y. (2002). Critical period for cross-modal plasticity in blind humans: a functional MRI study. Neuroimage, 16(2), 389-400.
Sadato, N., Okada, T., Kubota, K., & Yonekura, Y. (2004). Tactile discrimination activates the visual cortex of the recently blind naive to Braille: a functional magnetic resonance imaging study in humans. Neurosci Lett, 359(1-2), 49-52.
Sadato, N., Pascual-Leone, A., Grafman, J., Ibanez, V., Deiber, M. P., Dold, G., & Hallett, M. (1996). Activation of the primary visual cortex by Braille reading in blind subjects. Nature, 380(6574), 526-8.
Sampaio, E. (1994). [The early development of blind children: the pioneering work of Selma Fraiberg]. Psychiatr Enfant, 37(1), 29-47.
Sampaio, E., Maris, S., & Bach-y-Rita, P. (2001). Brain plasticity: 'visual' acuity of blind persons via the tongue. Brain Res, 908(2), 204-7.
Sampaio, E., & Philip, J. (1995). Influences of age at onset of blindness on Braille reading performances with left and right hands. Percept Mot Skills, 81(1), 131-41.
Schneider, G. E. (1973). Early lesions of superior colliculus: factors affecting the formation of abnormal retinal projections. Brain Behav Evol, 8(1), 73-109.
Sengpiel, F., Stawinski, P., & Bonhoeffer, T. (1999). Influence of experience on orientation maps in cat visual cortex. Nat Neurosci, 2(8), 727-32.
Sincich, L. C., & Horton, J. C. (2005). Input to V2 thin stripes arises from V1 cytochrome oxidase patches. J Neurosci, 25(44), 10087-93.
Spiers, H. J., & Maguire, E. A. (2006). Thoughts, behaviour, and brain dynamics during navigation in the real world. Neuroimage.
Sur, M. (1988). Visual projections induced into auditory thalamus and cortex: implications for thalamic and cortical information processing. Prog Brain Res, 75, 129-36.
Sur, M., Garraghty, P. E., & Roe, A. W. (1988). Experimentally induced visual projections into auditory thalamus and cortex. Science, 242(4884), 1437-41.
Sur, M., & Leamey, C. A. (2001). Development and plasticity of cortical areas and networks. Nat Rev Neurosci, 2(4), 251-62.
Sur, M., & Rubenstein, J. L. (2005). Patterning and plasticity of the cerebral cortex. Science, 310(5749), 805-10.
Toldi, J., Farkas, T., & Volgyi, B. (1994). Neonatal enucleation induces cross-modal changes in the barrel cortex of rat. A behavioural and electrophysiological study. Neurosci Lett, 167(1-2), 1-4.
Toldi, J., Rojik, I., & Feher, O. (1994). Neonatal monocular enucleation-induced cross-modal effects observed in the cortex of adult rat. Neuroscience, 62(1), 105-14.
Tomaiuolo, F., Ptito, M., Marzi, C.A., Paus, T., Ptito, A., (1997)Blindsight in hemespherectomised patients as revealed by spatial summation across the vertical meridian. Brain,120(5), 795-803
Voss, P., Gougoux, F., Lassonde, M., Zatorre, R. J., & Lepore, F. (2006). A positron emission tomography study during auditory localization by late-onset blind individuals. Neuroreport, 17(4), 383-8.
Wiesel, T. N., & Hubel, D. H. (1965). Comparison of the effects of unilateral and bilateral eye closure on cortical unit responses in kittens. J Neurophysiol, 28(6), 1029-40.
Weiskrantz, L., Warrington, E.K., Sanders, M.D., & Marshall, J. (1974) Visual capacity in the hemianopic field following a restrcited occipital ablation. Brain 97(4): 709-728.
Windrem, M. S., & Finlay, B. L. (1991). Thalamic ablations and neocortical development: alterations of cortical cytoarchitecture and cell number. Cereb Cortex, 1(3), 230-40.
Yu, H., Farley, B. J., Jin, D. Z., & Sur, M. (2005). The coordinated mapping of visual space and response features in visual cortex. Neuron, 47(2), 267-80.